My research focuses on unravelling the detailed organization of brain regions that are relatively expanded in humans: the association cortices. These same regions remarkably also serve advanced human functions such as language, episodic processes, and social cognition. My research has shown that association cortex contains parallel and closely interdigitated network architectures that require high-resolution methods performed at the level of individuals to be resolved (Braga and Buckner 2017; Braga et al. 2019). Further these closely neighboring parallel networks can be dissociated by different internally oriented processes (DiNicola et al. 2019).

I am currently an Instructor at Stanford University, supported by the NIH Pathway to Independence Award (K99), working with Josef Parvizi and Russ Poldrack. We are developing methods to combine individual-level whole-brain network mapping based on fMRI with intracranial recordings and stimulation in order to understand precisely how these distributed networks interact and compete during different forms of cognition.
As a Sir Henry Wellcome Postdoctoral Fellow at Harvard University, working with Randy Buckner, I focused on understanding the organization of distributed brain networks at the level of individuals.
Like a fingerprint, every brain is unique, containing its own arrangement of anatomical folds and functional zones. Because of this, brain imaging has to be performed at the level of individuals, preserving each individual’s unique circuitry, in order to reveal organizational details that may be key to understanding how cognitive functions are performed by the brain.
My research showed that focusing on the individual can reveal new principles of brain organization. In particular, we found that canonical association networks like the default network comprise multiple parallel distributed networks when resolved within individuals (Braga and Buckner 2017). The networks contain side-by-side regions throughout the brain, even within the same sulci when resolved at high resolution (Braga et al. 2019), and could be dissociated by episodic versus social cognitive processes (DiNicola et al. 2019).
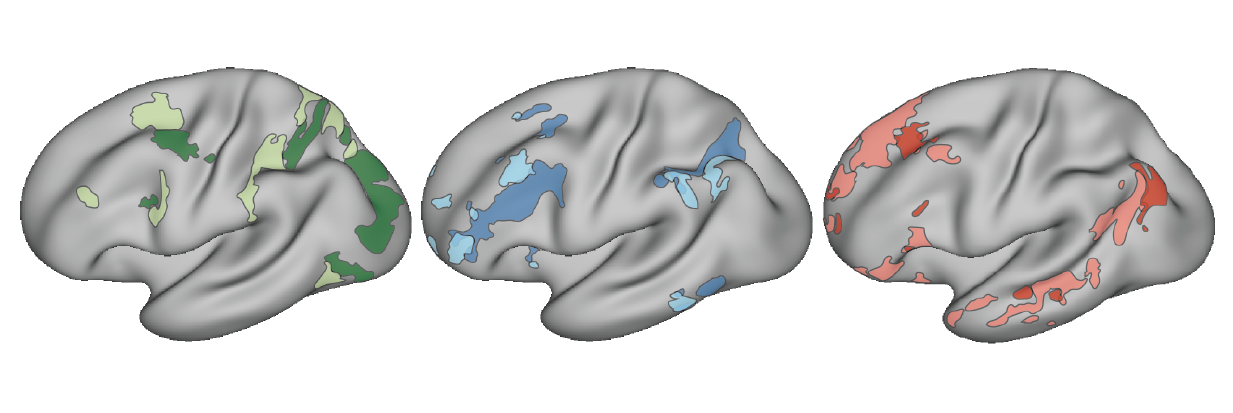
The group-defined dorsal attention (greens), frontoparietal control (blues), and default network (reds) were found to fractionate into parallel closely interdigitated networks when defined within individuals. Adapted from Braga & Buckner, 2017, Neuron.
This is especially interesting as these two processes, mental projection in time and thinking about what others are thinking, are both internally oriented and involve content not borne from your current sensory environment: in one case a mentally constructed scene and event, and in the other the consideration of someone else’s beliefs, desires, intent, etc. These different private mental phenomena map onto separate networks that share a similar broad structure, containing side-by-side regions in multiple parts of the brain.
My PhD research was supervised by Dr. Robert Leech and Prof. Richard Wise, and focused on the brain networks that are recruited when we pay attention to different sounds. We found evidence that separable distributed systems serve the orienting of attention to vision and sound (Braga et al. 2013), and that these systems interact during natural listening conditions (Braga et al. 2016; Braga et al. 2017).
We also investigated the organization of distributed networks in group-averaged data and showed that multiple networks can be detected in the same focal regions of association cortex (Leech et al. 2012; Braga et al. 2013), suggesting that a finer-scale functional architecture exists that lies beyond the resolution of group-averaging. This spurred me to begin an exploration of functional anatomy at the level of individuals using repeated sampling approaches at Harvard.

Separable networks observed during top-down attention to auditory and visual modalities. Adapted from Braga et al., 2013, Neuroimage.

Eye movements during auditory attention: Listening to sounds in the left and right ears led to differences in natural gaze position. Adapted from Braga et al., 2016, Frontiers in Human Brain Mapping.
PUBLICATIONS
1. R.M. Braga, K.R.A. Van Dijk, J.R. Polimeni, M.C. Eldaief & R.L. Buckner (2019) Parallel Distributed Networks Resolved at High Resolution Reveal Close Juxtaposition of Distinct Regions. Journal of Neurophysiology, 121: 1513-1534.
2. L.M. DiNicola, R.M. Braga, R.L. Buckner (2019) Parallel Distributed Networks Dissociate Episodic and Social Functions Within the Individual. bioRxiv, https://doi.org/10.1101/733048.
3. S. Kamourieh, R.M. Braga, R. Leech, A. Mehta & R. Wise (2018) Speech Registration in Symptomatic Memory Impairment. Frontiers in Aging Neuroscience, 10: 201.
4. R.M. Braga & R.L. Buckner (2017) Parallel Interdigitated Distributed Networks within the Individual Estimated by Intrinsic Functional Connectivity. Neuron, 95: 457-471.
5. R.M. Braga, P.J. Hellyer, R.J.S. Wise & R. Leech (2017) Auditory and visual connectivity gradients in frontoparietal cortex. Human Brain Mapping, 38: 255-270.
6. R.P. Monti, R. Lorenz, R.M. Braga, C. Anagnostopoulos, R. Leech & G. Montana (2017) Real-time estimation of dynamic functional connectivity networks. Human Brain Mapping, 38: 202-220.
7. R.M. Braga, R.Z. Fu, B.M. Seemungal, R.J.S. Wise & R. Leech (2016) Eye movements during auditory attention predict individual differences in dorsal attention network activity. Frontiers in Human Neuroscience, 10: 164.
8. R.M. Braga & R. Leech (2015) Echoes of the Brain: Local-Scale Representation of Whole-Brain Functional Networks within Transmodal Cortex. The Neuroscientist, 21: 540-551.
9. R.M. Braga, Roze E, Ball G, Merchant N, Tusor N, Arichi T, Edwards D, Rueckert D & Counsell SJ (2015) Development of the Corticospinal and Callosal Tracts from Extremely Premature Birth up to 2 Years of Age. PLoS One, 10 (5) e0125681.
10. S. Kamourieh, R.M. Braga, R. Leech, R.D. Newbould, P. Malhotra, R.J.S. Wise (2015) Neural systems involved when attending to a speaker. Cerebral Cortex, 25: 4284-4298.
11. S.R. Jilka, G. Scott, T. Ham, A. Pickering, V. Bonnelle, R.M. Braga, R. Leech, D.J. Sharp (2014) Damage to the salience network and interactions with the default mode network. The Journal of Neuroscience, 34: 10798-10807.
12. R.M. Braga, D.J. Sharp, C. Leeson, R.J.S. Wise & R. Leech (2013) Echoes of the brain within default mode, association and heteromodal cortices. The Journal of Neuroscience, 33: 14031-14039.
13. R.M. Braga, L.R. Wilson, R.J.S. Wise, D.J. Sharp & R. Leech (2013) Separable networks for top down attention to auditory nonspatial and visuospatial modalities. NeuroImage, 74: 77-86.
14. R. Wise & R.M. Braga(2013) Default mode network: the seat of literary creativity? Trends in Cognitive Sciences, 18: 116-117.
15. R. Leech, R.M. Braga & D.J. Sharp (2012) Echoes of the brain within the posterior cingulate cortex. The Journal of Neuroscience, 32: 215-222.
16. E. Rose, P. Harris, G. Ball, L.Z. Elorza, R.M. Braga, J.M. Allsop, N. Merchant, E. Porter, T. Arichi, A.D. Edwards, M.A. Rutherford, F.M. Cowan & S.J. Counsell (2012) Tractography of the corticospinal tracts in infants with focal perinatal injury: comparison with normal controls and to motor development. Neuroradiology, 54: 507-516.
17. M.A.A. van der Meer, Z. Richmond, R.M. Braga, P.A. Dudchenko & E.R. Wood (2010) Evidence for the use of an internal sense of direction in homing. Behavioral Neuroscience, 124: 164-169.